
Диана Вальштад. Аллелопатия

У меня в аквариуме объемом 55 галлонов (~200 л) с садовой почвой и хорошим освещением некоторые растения растут хорошо, а другие плохо. Пять анубиасов нана (Anubias Nana) и четыре эхинодоруса сердцелистных (Echinodorus cordifolius) чувствуют себя как дома, а вот с гигрофилой многосеменной (Hygrophila polysprema) и хоттонией инфлата (Hottonia inflata) дела не очень. Листья гигрофилы скручиваются и опадают, а у хоттонии на некоторых листьях образовывается коричневый налет.
Может это температура влияет на появление коричневого налета? Я в какой-то книге читал, что для этого растения температура должна быть в пределах 64 - 73°F (18 - 23°C), а у меня 77°F (25°С)
О.
Я бы не слишком переживала, что некоторые растения не очень хорошо себя чувствуют, при условии, что другие процветают. Конечно же некоторые виды растений будут расти хуже других, так как данные условия содержания для них не подходят Ваша хоттония любит прохладную воду, а гигрофиле, может, нужно больше CO2.
А может быть и такое, что другие растения вашего аквариума выделяют химические вещества («аллелохимикаты»), которые угнетают и гигрофилу и хоттонию. Явление аллелопатии среди растений может объяснить многие случаи, когда конкретный вид не растет в конкретном домашнем аквариуме. Например, эхинодорусы амазонские (Echinodorus amazonicus), анубиасы нана (Anubias Nana), лимнофилы и некоторые криптокорины прекрасно уживаются вместе в моем аквариуме на 50 гал. (~200 л). Однако в этом же аквариуме я не могу вырастить валлиснерию. Причем она прекрасно чувствует себя в других моих аквариумах. Я принимаю факт аллелопатии среди растений как естественный и неизбежный.
***
Теоретически, аллелопатия - это производство и высвобождение организмами в окружающую среду химических («аллелохимических») веществ, которые воздействуют на другие организмы. Хотя некоторые животные также производят защитные химические вещества, но обычно аллелохимические вещества вырабатываются растениями и другими неподвижными организмами, и наиболее вероятным воздействием этих веществ на целевые объекты является их подавление. Ведь растения не могут защититься от врага ни габаритами, ни скоростью, ни силой. От болезней и поедания травоядными им остается использовать в основном только химическую защиту [1, 2].
И, похоже, растения серьезно вложились в создание химической защиты. Ведь аллелохимикаты - это не отходы жизнедеятельности, но вырабатываются специально, причем при значительных энергозатратах. Растения активно отбирают ценные аминокислоты фенилаланин и тирозин от синтеза белка для метаболического пути фенилпропаноидов, чтобы вырабатывать фенольные кислоты, дубильные вещества, флавоноиды, стильбены и лигнины:
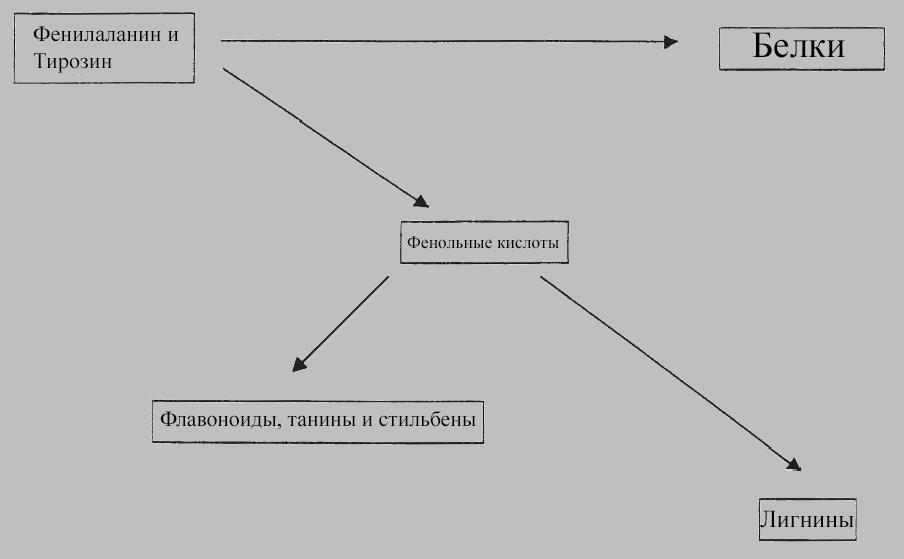
Фенольные аллелохимикаты являются продуктами метаболического пути фенилпропаноидов.
Многие из этих соединений являются аллелопатическими. Химическая структура трех распространенных фенольных аллелохимикатов показана на рисунке:
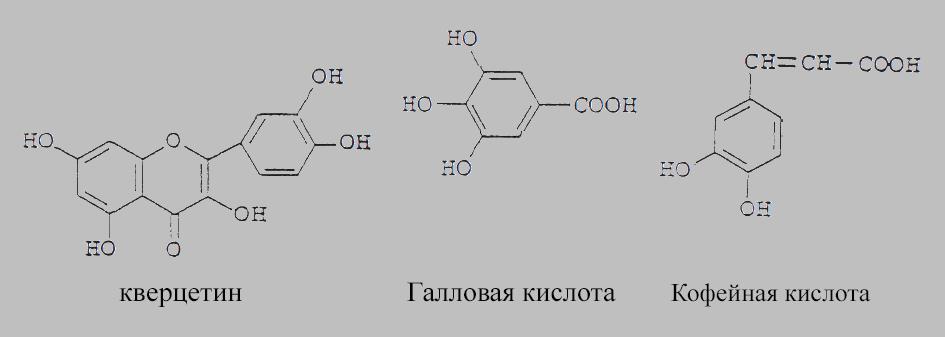
Химическая структура нескольких фенольных аллелохимикатов.
Галловая и кофейная кислоты являются фенольными кислотами, а кверцетин - флавоноидом.
Поскольку аллелохимические вещества могут угнетающе действовать и на растение-хозяина, то эти вещества надо как-то надлежащим образом хранить. Поэтому химзащита с помощью аллелохимических веществ требует больше усилий, чем просто их производство. Но, похоже, это того стоит, ведь 10-ти процентное метаболическое вложение растений в аллелохимические вещества может предотвратить 90 процентную потерю от травоядных.
Смею предположить, что аллелопатия в водной среде будет иметь много случайных побочных эффектов. Большинство аллелохимикатов растворимы в воде, поэтому несмотря на то, что целью растения является организм-мишень, они, хочешь не хочешь, но будут влиять и на другие окружающие организмы. А в закрытой среде, какой является аквариум, аллелохимические вещества имеют свойство накапливаться, а их эффект усиливаться.
Аллелопатия водных растений
Водные растения содержат разнообразные аллелохимические вещества, основной функцией которых является защита растения от поедания рыбой и насекомыми или уничтожения болезнями.
Прим. аллелохимические вещества могут иметь и другие функции. Например, два флавоноида, апигенин и лютеолин, являются аллелопатическими, но они также могут обеспечивать защиту от вредного ультрафиолетового излучения, что полезно для роста в воздухе некоторых водных растений [3]. А кофеиновая и хлорогеновая кислоты, по-видимому, действуют как хелаторы для корневого поглощения железа некоторыми наземными растениями [4, 5].
В целом, водные растения считаются более устойчивыми к болезням и поеданию травоядными животными, чем наземные растения [6].
Об аллелопатическом поведении сообщалось у 97 видов водных растений [7]. Действительно, когда исследователи тестировали экстракты из 17 различных видов водных растений, все 17 экстрактов подавляли ростки ряски или салата:
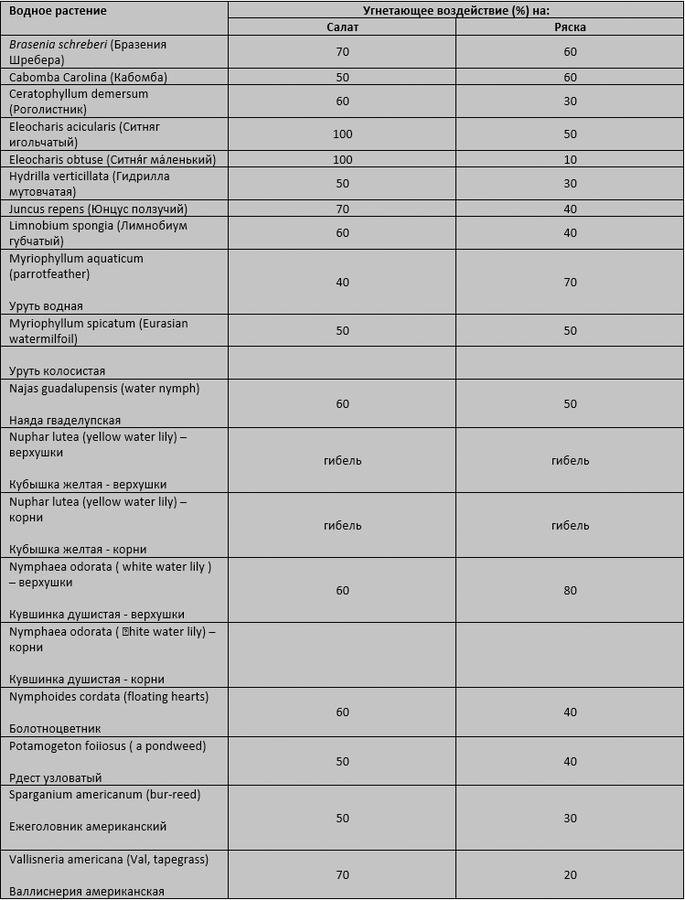
Токсичность экстрактов водных растений. Елакович и Вутен (Elakovich and Wooten) (8,9]. Процент подавления ростков салата, выращенных в чашках Петри, содержащих экстракты растений, вычислялся путем измерения длины их корней и сравнивался с контрольными (выращенных в чашках Петри без экстрактов растений). Вычисление подавления роста ряски представляет собой количество новых ветвей в питательных средах с растительными экстрактами в сравнении с контрольными (ряска без растительных экстрактов). Все растительные экстракты, за исключением водяных лилий, готовили из цельных растений. Двести граммов свежего растительного вещества каждого вида измельчали и тщательно смешивали с 200 мл дистиллированной воды и охлаждали в течение 1 - 3 дней. Экстракты фильтровали и затем разводили (1 : 5) в питательной среде салата и ряски.
Эти два исследования проводились в том числе и над такими распространенными аквариумными растениями, как кабомба, роголистник и валлиснерия. Растением с самым сильным подавляющим действием оказалась кубышка жёлтая (Nuphar lutea), которая стала причиной гибели (а не просто угнетения) ростков салата и ряски.
Аллелохимикаты, выделенные из водных растений (Таблица III-2), подавляют различные организмы (Таблица III-3):

Таблица III-2. Аллелохимикаты, найденные в водных растениях.
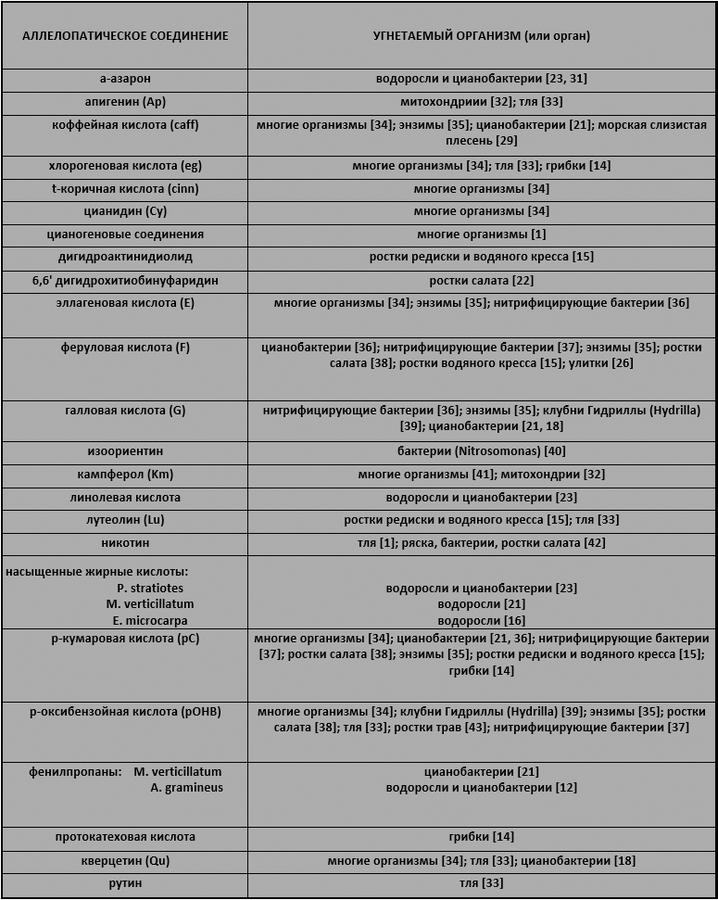
Таблица III-3. Аллелопатия соединений, обнаруженных в водных растениях. Сокращения следуют общеизвестным аллелопатическим фенолам. Caff, cinn, E, F, G, pC, pOHB, протокатехиновая кислота, S, Sy и V представляют собой простые фенольные кислоты и фенилпропаны, в то время как Ap, Cy, Km, Lu и Qu представляют собой флавоноиды. Линолевая кислота представляет собой жирную кислоту С18. Недавно выделенные и идентифицированные фенилпропаны, насыщенные кислородом жирные кислоты, стерины и дубильные вещества описаны в ссылках.
1. Фенольные соединения, как аллелохимикаты водных растений
Важную роль в аллелопатии водных растений играют фенольные соединения (а не алкалоиды и т. д.).
Прим. Алкалоиды, такие как никотин, дигитоксин, стрихнин, морфин и кураре - являются хорошо известными аллелохимическими веществами наземных растений [1]. Различные алкалоиды были обнаружены в кувшинках [17]. Однако в других водных растениях алкалоидов по-видимому мало. Было обнаружено, что пятнадцать видов подводных растений содержат менее 0,06% алкалоидов [44].
Это связано с тем, что фенольные соединения являются частью метаболизма фенилпропаноидов для синтеза лигнина, который обеспечивает поддержку наземным растениям, в том числе деревьям, давая им возможность стоять в вертикальном положении. В ходе эволюции, когда наземные растения перешли в воду, чтобы стать водными растениями, они потеряли свою потребность в лигнине, потому что сама плотность воды обеспечивала им необходимую поддержку. Таким образом, содержание лигнина постепенно снижалось.
Прим. McClure [11] предоставляет филогенетические доказательства постепенного сокращения содержания лигнина, которое происходило вместе с развитием водных растений. Ванилин и сирингальдегид являются характерными фенольными предшественниками лигнина. Более примитивные виды рясковых (Lemnaceae) (например, Spirodela intermedia, S. polyrhiza и S. oligorhiza) содержат эти фенолы, тогда как более развитые виды рясковых (например, Lemna minor, L. gibba и L. trisulca) не содержат.
В настоящее время большинство подводных растений имеют малое содержание не нужного более лигнина, однако они все еще содержат его прекурсоры[45,46].
Тот факт, что прекурсоры лигнина оказывают не такое жесткое подавляющее действие на различные организмы, оказался для водных растений удачным . Поскольку фенилпропаноидный путь уже имелся, то для производства аллелохимикатов водным растениям не нужно было создавать совершенно новый метаболический путь . В ходе эволюции уже происходили спонтанные мутации, увеличивающие подавляющие свойства производимых фенольных соединений. Действительно, один исследователь (31) продемонстрировал, как простые химические изменения обычных фенольных кислот могут резко повлиять на их угнетающее воздействие на водоросли.
Чем выше содержание фенолов в водном растении, тем меньше вероятность его поедания травоядными [47,48]. Растения, содержащие более 6% фенольных соединений, считаются неперевариваемыми и имеют небольшую пищевую ценность. (Сельскохозяйственные кормовые культуры, выведенные для лучшей поедаемости, содержат менее 2 - 3% фенольных соединений.) Содержание фенолов в водных растениях в среднем составляет около 6% в диапазоне от 0,8% у элодеи густолиственной (Elodea densa) до 15% у кабомбы каролинской (Cabomba caroliniana) [45].
Растения, обладающие фенольными веществами, могут оказывать аллелохимическое воздействие друг на друга. Например, было обнаружено, что виды растений (кувшинка душистая (Nymphaea odorata), бразения шребера (Brasenia schreberi) и кабомба каролинская (Cabomba caroliniana)), содержащие самые высокие уровни фенольных соединений [45,46], сильнее всех подавляют ряску [8].
Фенольные соединения оказывают угнетающее воздействие на различные организмы из-за их способности к выведению из строя белков. Так, например, в кожевенной промышленности процесс дубления кожи основан на способности растительных полифенолов, таких как дубильные вещества, деактивировать и полимеризовать белки. В живущем растении эти же дубильные вещества препятствуют поеданию его насекомыми, повреждая белки кишечника последнего.
Фенольные кислоты обнаруживаются в очень высоких концентрациях в специализированных «фенольных клетках». У водного гиацинта фенольные клетки, вкупе с обычными клетками, находятся главным образом в субэпиденальной ткани обеих поверхностей листьев [51]. Фенольные кислоты в этих клетках находятся в очень высоких концентрациях - около 1000 ppm - и состоят из хлорогеновой, протокатехиновой, ванильной и p-кумаровой кислот [14]. Считается, что фенольные клетки играют роль в устойчивости водного гиацинта к грибку, ответственному за болезнь «пятнистости листьев» [52].
2. Выделение растениями аллелохимических веществ
Выделяют ли водные растения аллелохимические вещества в воду? Если они остаются крепко запертыми внутри растения, то их влияние на окружающую среду аквариума - водоросли, бактерии или другие растения будет ограничено.
Наземные растения часто выделяют аллелохимические вещества в окружающую среду (36). Например, было обнаружено, что корни молодых деревьев папайи выделяют аллелохимический бензилизотиоцианат из расчета 2 мкг с дерева в день (53). Чапараль выделяет из листьев различные водорастворимые фенольные кислоты, такие же, как у водных растений. Эти вещества смываются дождевой водой в почву, где они предотвращают прорастание и рост конкурирующих растений [43].
Живые водные растения, теряют большое количество аллелохимикатов, вследствие их текучести, а погибшие растения теряют еще больше. Считается, что живые погруженные водные растения выделяют около 4% растворенного органического углерода (РОУ) от общего зафиксированного годового количества, а отмершие - 40%. Бактерии перерабатывают большую часть этого РОУ в гуминовые вещества. Кроме того, водные растения постоянно заменяют старые, разлагающиеся листья новыми. Например, сообщается, что кувшинка Nymphaea odorata, произрастающая на юге США, 7 раз в год полностью меняет листья. Такой обильный оборот биомассы влечет за собой повышенный выброс в воду аллелохимических веществ.
Действительно, в питательных средах водных растений были обнаружены аллелохимикаты. При выращивании ряски в стерильных питательных средах, в них (в средах) быстро обнаруживаются коричные кислоты, а через несколько дней обнаруживаются флавоноиды [11]. И, как сообщается, перистолистник бразильский (Myriophyllum brasiliense) выделяет в питательную среду небольшое количество своих аллелопатических полифенолов [18]. Выделение фенольной кислоты урутью колосистой (Myriophyllum spicatum) за 10 дней составило 2 - 4 мг/г сухого веса растительного вещества [20]. Несколько аллелохимических веществ были обнаружены в прудовой воде, в которой росла Eleocharis microcarpa [16].
Хотя большая часть РОУ, выделяемого водными растениями, быстро перерабатывается бактериями, всегда есть часть, которая не разлагается. Например, большая часть РОУ, высвобождаемого Scripus subterminalis, перерабатывается бактериями в течение 3 дней, но 5 - 10% остается нетронутыми и через 40 дней [56]. Этот оставшийся РОУ содержит фенольные соединения (как синтезированные аллелохимические, так и гуминовые вещества), потому что они по своей природе устойчивы к бактериальному разложению. (Было установлено, что скорость разложения различных компонентов растительных остатков через один год составляет 99% для сахара, 90% для гемицеллюлозы, 75% для целлюлозы, 50% для лигнина, 25% для восков и только 10% для фенольных соединений [57]).
3. Тонкая сущность аллелопатии водных растений
Большинство растительных аллелохимикатов обладают слабыми ингибирующими свойствами. Из-за этого ученым трудно доказать аллелопатию.
Очень высока вероятность того, что аллелопатия растений является результатом комбинированного действия многих слабодействующих химических веществ. Таким образом, ученые продолжают искать более точные доказательства воздействия специфических и сильнодействующих фитотоксинов, хотя на самом деле угнетающий эффект на растения может заключаться в совместном действии большого количества слабых токсических соединений [43].
В самом деле, аллелохимическое вещество может оказывать больший угнетающий эффект, когда комбинируется с другими аллелохимикатами, чем когда тестируется отдельно (т.е. имеет место «взаимоусиливающий эффект») [59]. Например, две не слишком сильные фенольные кислоты (галловая кислота и кофейная кислота) подавляли рост сине-зеленых водорослей в 6 раз сильнее, когда их смешивали вместе, чем когда их тестировали отдельно [21]. Это важный вывод, потому что низкая активность многих фенольных аллелохимикатов говорит о том, что они могут действовать слабо или вообще не действовать за пределами лаборатории. Тем не менее, если присутствует много аллелохимикатов (а это так на самом деле), и они действуют синергетически, то аллелопатия возможна .
Аллелопатия водных растений не впечатляет. Она едва различима. Тем не менее, все водные растения постоянно производят большое и разнообразное количество защитных соединений, которые слегка угнетают все организмы. Вполне вероятно, что в водных экосистемах эти аллелохимические вещества могут оказывать едва заметное и нераспознаваемое воздействие на растения, бактерии, водоросли и беспозвоночных .
4. Водные растения против водорослей
Специалисты по водной ботанике обнаружили, что в озерах с интенсивным ростом растений, рост водорослей часто снижается [20]. Конечно, отчасти это видимое угнетение может быть связано с конкуренцией растений и водорослей за свет и питательные вещества. Тем не менее, некоторое ингибирование может быть следствием воздействия аллелохимических веществ растений. Также оно может быть связано с гуминовыми веществами, которые могут угнетать организмы [61]. Гуминовые вещества, являющиеся фенольными соединениями, образуются в результате разложения (а не синтеза) растительных фенольных соединений.
Один исследователь [62] отслеживал рост водорослей в зависимости от концентрации фенольных соединений (в основном гуминовых веществ) в 6 испанских прудах в течение двухлетнего периода. Из-за сезонных паводков концентрация фенолов в прудах варьировалась от 4 до 26 мг/л. Когда концентрация была на уровне 10 мг/л или выше, а уровни питательных веществ были низкими, рост водорослей уменьшался. В исследовании с использованием фенольных экстрактов из урути колосистой (Myriophyllum spicatum) [19] концентрация фенольных соединений 10 мг/л оказывала умеренный ингибирующий эффект на водоросли и цианобактерии.
Вероятно, не только одни аллелохимикаты и неспецифичные гуминовые кислоты действуют на рост водорослей. Chara globularis производит два серосодержащих соединения, дитиолан и тритиан, которые, как было установлено, сильно тормозят фотосинтез водорослей [13]. В другом исследовании 33 из 43 различных оксигенированных жирных кислот, обнаруженных в прудовой воде, содержащей Eleocharis microcarpa, в искусственных условиях подавляли рост сине-зеленых [16].
В таблице показано подавление роста различных водорослей аллелохимическими веществами полупогруженного растения рогоза широколистного (Typha latifolia):
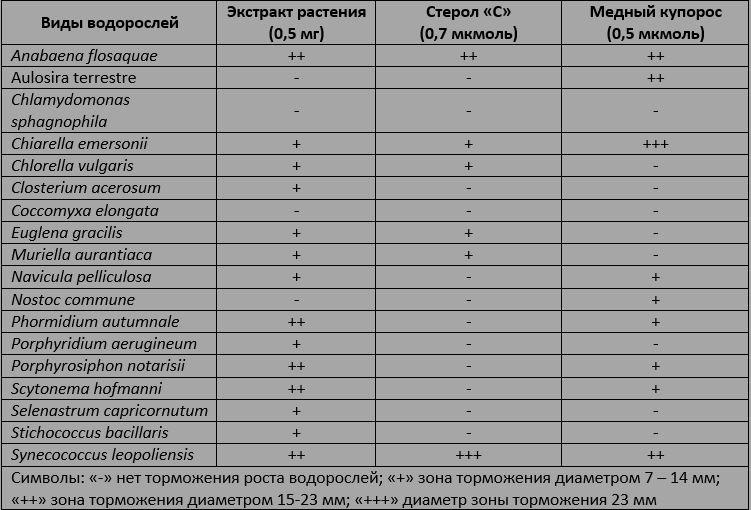
Сравнительная таблица подавления роста водорослей рогозом широколиственным (Typha latifolia) и медным купоросом. Растительный экстракт сделан на этиловом эфире. Стерол "C" - стигмаст-4-ен-3-он, 6-дион. Биологический анализ проводили в чашках Петри, содержащих питательный агар, засеянный экспоненциально растущими водорослями. Растворы ацетона, содержащие известные количества химических веществ, высушивали на фильтровальных дисках, которые затем добавляли в чашки Петри. Чашки инкубировали на свету, пока рост водорослей не становился видимым. Подавление роста проявлялось в виде прозрачных зон вокруг фильтровальных дисков.
В таблице показано сравнение действия аллелохимических веществ растения с эффектом медного купороса. Два вида сине-зеленых водорослей (Anabaena Flosaquae и Synechococcus leopoliensis) оказались достаточно чувствительны как к неочищенному растительному экстракту, так и к стеролу.
Хотя большинство аллелохимикатов водных растений слабо подавляют водоросли, некоторые все же обладают более сильными угнетающими свойствами. Изучая поглощение питательных веществ из загрязненных вод, у исследователей [18] возникло подозрение, что Myriophyilum brasiliense выделяет ингибирующие вещества против соседних сине-зеленых водорослей. Используя осторожные методы экстракции, они смогли выделить из растения два очень сильных полифенола: Теллимаграндин II и 1-дезгаллолейгениин.
Доминирование урути колосистой (Myriophyllum spicatum) в озерах Северной Америки может быть связано с ее фенольными свойствами. Описывалось, что фенольные соединения растения в концентрации 10 мг/л полностью подавляют рост сине-зеленых водорослей. Зеленые водоросли подавлялись концентрацией 20 мг/л [19]. Теллимаграндин II, который был впервые обнаружен у наземного многолетнего растения теллима крупноцветковая (Tellima grandiflora), а затем у других представителей отряда Rosales [63], был обнаружен в высоких концентрациях у урути колосистой (M. spicatum) [20]. Исследователи подсчитали, что если бы уруть колосистая (M. spicatum) высвободила всего лишь 1% своего теллимаграндина II, то этого выброса было бы достаточно, чтобы серьезно повлиять как на эпифитные (прикрепленные к растению), так и на планктонные (взвешенные) водоросли [64].

Уруть колосистая (Myriophyllum spicatum). M. spicatum, по-видимому, вырабатывает аллелохимические вещества против различных организмов (ряски, сине-зеленых водорослей, личинок комаров и водного растения наяда морская (Najas marina). Она выделяет довольно сильнодействующий аллелохимический Tellimagrandin II, который может защищать ее от водорослей.
Недавно группа исследователей методично отслеживала несколько водных растений на наличие аллелохимических веществ против водорослей. Из Аира злакового (Acorus gramineus) было выделено семь различных фенольных кислот, в том числе те, которые подавляли некоторые виды водорослей и цианобактерий с токсичностью, сравнимой с медным купоросом [12]. У водяного салата (Pistia stratiotes) исследователи также обнаружили ассортимент аллелохимических веществ - стеролы, полипренолы, жирные кислоты и α-азарон. Наиболее сильнодействующим ингибирующим соединением был α-азарон, который подавлял рост 14 из 19 протестированных видов водорослей [31].
Хотя вышеприведенные исследования показывают, что растения содержат небольшие количества сильнодействующих ингибиторов водорослей, таких как α-азарон и tellimagrandin II, многие водные растения могут не производить эти соединения в количествах, достаточных для сдерживания роста водорослей в природе (или в наших аквариумах). По большей части аллелопатия водных растений, вероятно, связана с общим количеством (~6% от сухого веса растений) всего разнообразия фенольных кислот.
5. Водные растения против бактерий и беспозвоночных
Поскольку аллелохимические вещества зачастую являются не особо разборчивыми ингибиторами, то водные растения могут подавлять и бактерий. Например, экстракты бразении шребера (Brasenia schreberi ) были протестированы против 9 видов бактерий, как грамотрицательных, так и грамположительных: все 9 видов подавлялись различными объемами растительного экстракта [66]. Экстракты кувшинки Nymphaea tuberosa показали высокое противомикробное действие в отношении нескольких видов бактерий (67]. Аллелохимические вещества, ответственные за ингибирование, были идентифицированы как дубильная кислота, галловая кислота и этилгаллат. Все они являются распространенными фенольными соединениями, встречающимися во многих водных растениях. Более того, некоторые исследования показывают, что аллелохимия, вырабатываемая водными растениями, подавляет цианобактерии («сине-зеленые водоросли»). (Это означает, что другие бактерии также могут подавляться.)
Водные растения, по-видимому, выделяют в воду химические вещества, которые отпугивают беспозвоночных. Так, в экспериментальных резервуарах дафния уплывала от элодеи, урути и нителлы чаще, чем от пластиковых растений в контрольных резервуарах [68]. Еще один исследователь [69] продемонстрировал, что экстракты урути колосистой (Myriophyllum spicatum ) отпугивают мошек и личинок комаров. Аллелопатия может объяснить уменьшение популяции, комаров, мошек и дафний, которое ученые наблюдали в стоячих озерах с интенсивным ростом растений.
Улитки избегают поедания здоровых листьев водных растений, но едят отмершие или больные [70,71]. Например, когда обыкновенным литоринам предлагали свежесобранные листья солоноводной Spartina altemiflora, они предпочитали мертвые (но неповрежденные) листья здоровым листьям в соотношении около трех к одному. Считалось, что разницу в предпочтениях объясняет более низкое содержание феруловой кислоты в мертвых листьях. (Из отмерших листьев феруловая кислота попросту вытекает, делая их менее токсичными.)
В.
Выращивание валлиснерии гигантской оказалась для меня большой проблемой, хотя предполагается, что это относительно простое растение. После первоначального бурного роста, лихо закрутившись под поверхностью воды, растения, кажется, начинают уменьшаться.
Другая проблема, с которой я столкнулся, связана с улитками Мистериями. Несмотря на то, что улитки время от времени жевали молодые ростки гигрофилы многосеменной (H. polysperma), они не наносили ей непоправимого ущерба. А вот валлиснерия гигантская, похоже, стала у них любимой едой. Они едят ее, пока она почти не заканчивается. Те же улитки, помещенные в аквариум со спиральной валлиснерией (V. spiralis), возвращаются к питанию исключительно водорослями.
Почему они едят гигантскую валлиснерию и не едят спиральную? Никакие другие растения в 70 галл. (~280 л) аквариуме не подвергаются такому нападению. У других любителей подводного сада был подобный опыт? Есть ли другие растения, для которых улитки Мистерии могут быть опасны?
О.
Ваш вопрос о валлиснерии гигантской (Vallisneria gigantea) и улитках Мистериях наиболее интересен. Хотя у меня нет практической информации об улитках Мистериях, поедающих растения, но все, что я читала в научной литературе, говорит о том, что большинство видов улиток полезны растениям, так как очищают листья и едят только мертвые или гибнущие растения. [Все здоровые водные растения содержат защитные химические вещества (аллелохимикаты), которые отпугивают улиток и других травоядных, но как только растительная ткань начинает распадаться, эти токсичные химические вещества вымываются. Только тогда улитки поедают растение.] Поскольку вы описали проблему с V. gigantea, то может быть, она гибнет и улитки просто потребляют умирающее растение, а здоровую спиральную валлиснерию не трогают?
6. Химическая война между водными растениями
Водные растения часто лучше растут в одиночку, чем в паре с другими видами [67]. Помимо защиты себя от поедания травоядными, водные растения также синтезируют аллелохимические вещества, которые делают их более конкурентоспособными в их непосредственной среде обитания. Другими словами, они хотят отравить соседей и захватить их территорию.
а) Аллелопатия в грунте
Выделение аллелохимических веществ в субстрат было убедительно доказано для Eleocharis coloradoensis. В природе это крошечное растение, выделяя аллелохимикаты в субстрат, могло уничтожить густые насаждения крупных прудовых растений.
В серии экспериментов исследователи [72] впервые показали, что прудовые растения рдест узловатый (Potamogeton nodosus) и особенно рдест гребенчатый (P. pectinatus) плохо размножались, когда их клубни были посажены в почву, в которой рос Eleocharis coloradoensis. Впрочем, поскольку растения вместе росли в одних и тех же аквариумах , снижение роста прудовых растений могло быть связано с конкуренцией за питательные вещества или возможными изменениями качества воды элеохарисом.
Eleocharis coloradoensis. Это маленькое (5 - 7 см) почвопокровное растение, найденное в западной части США, по-видимому, может успешно конкурировать с гораздо более крупными растениями, выделяя в грунт отравляющие вещества. Хотя предполагается, что аллелопатия присуща и другим элеохарисам, но в отношении E. coloradoensis она убедительно доказана.
Таким образом, те же исследователи приступили к более точному эксперименту, где растения выращивались в отдельных аквариумах. Eleocharis coloradoensis в течение 3-х месяцев выращивались в одном контейнере, а прудовые растения росли в других, нижестоящих контейнерах. Днища контейнеров соединялись между собой пластиковыми шлангами. Вода, движимая силой тяжести, медленно просачивалась вниз через почву, где рос элеохарис, и затем проходила сквозь почву культур прудовых растений. Контрольным резервуаром являлась такая же конструкция, но без элеохариса. Вода из почвы, в которой рос элеохарис повлияла на рост прудовых растений, снизив его почти на половину. У испытуемых растений также наблюдался очевидный хлороз. Исследователи были внимательны к тому, чтобы содержание питательных веществ в фильтрате элеохариса было таким же как и у чистой почвы, что указывает на то, что недостаток питательных веществ не являлся причиной плохого роста прудовых растений.
Однако, даже эти эксперименты не доказали окончательно, что элеохарис обладал аллелопатическими свойствами. Бактерии и другие микробы в корневой зоне могут усиливать или разрушать аллелохимические вещества, выделяемые корнями растений. Почвенный гумус и глина могут поглощать аллелохимические вещества и уменьшать их подавляющее действие [73, 74]. Все эти факторы могут влиять на аллелопатическую активность в иле и почве. Вот если бы аллелопатические свойства элеохариса оставались бы в отсутствие бактерий и частиц почвы, тогда их можно было бы отнести непосредственно к аллохимическим веществам этого растения. Когда в более поздних исследованиях Eleocharis coloradoensis рос в песке и стерильной питательной среде, то его корневые выделения все также обладали ингибирующими свойствами (по отношению к рдесту гребенчатому P. pectinatus и гидрилле мутовчатой Hydrilla vercillata) [75]. Эти последние эксперименты дают убедительные доказательства того, что элеохарис выделяет ингибирующие аллелохимические вещества в субстрат.
Несмотря на то, что элеохарис содержит несколько известных ингибирующих соединений, считается, что его аллелопатия обусловлена главным образом дигидроактинидиолидом [15].
б) Аллелопатия в воде
Аллелопатия встречается также в толще воды и может быть довольно специфичной. Например, исследователи [77] высадили двадцать наяд поодиночке и попарно с 20 растениями другого вида в больших (200 литровых) контейнерах, содержащих песчано-суглинистую почву. Напарниками наядам стали три других вида: рдест блестящий (Potamogeton lucens), камыш прибрежный (Scirpus litoralis) и уруть колосистая (Myriophyllum spicatum). В течение 2-месячного периода наяды росли при достаточном пространстве, чтобы растения не были ограничены ни свободным местом, ни питательными веществами. Результаты показали, что с рдестом блестящим и с камышом прибрежным наяда росла так же хорошо, как и в одиночку. Однако в паре с урутью колосистой рост наяды замедлился вдвое.
В другом эксперименте воду из чистых культур урути колосистой (Myriophyllum spicatum) каждую неделю добавляли в контейнеры с наядой. В течение лета рост наяды в такой воде составлял менее 1/3 его роста в обычной водопроводной воде. И снова исследователи осторожно подошли к эксперименту, чтобы показать, что истощение питательных веществ не было причиной плохого роста наяды. Также было показано, что и рост урути колосистой подавлялся наядой. Результаты могут объяснить, почему наяда морская (Najas marina) и уруть колосистая (Myriophyllum spicatum) обычно не растут вместе в естественных водоемах (Израиля).
По-видимому, и гидрилла с роголистником иногда плохо уживаются в природе. Исследователи [78] пытались найти причину, по которой всего несколько побегов гидрилл, попадающих в индийские пруды и водохранилища, могли быстро и полностью уничтожить заросли роголистника. Так в одном эксперименте Ceratophyllum demersum и C. muricatum растили в резервуарах с садовой землей - в одном резервуаре отдельно, а в другом вместе с гидриллой мутовчатой (Hydrilla verticillata). Растения были разделены проволочной сеткой, чтобы они не находились в прямой конкуренции - они просто росли в одной воде. Результат оказался драматичным. Первоначально виды Hydrill a и Ceratophyllum хорошо росли вместе, но через 30 дней Ceratophyllum побледнел и постепенно распался. Через 70 дней роголистник канул в Лету, а гидрилла хорошо росла на всем доступном пространстве. Контрольные растения (Ceratophyllum demersum и C. muricatum, живущие без гидриллы) были здоровыми и хорошо росли.
7. Химическая защита от инфекции
Хотя все растения содержат большое количество фенольных кислот, некоторые фенольные кислоты могут вырабатываться в ответ на инфекцию [79]. Например, слизистая плесень Labyrinthula zosterae в 1930-х годах опустошала североатлантические плантации морских водорослей Zostera marina. Когда исследователи [80] преднамеренно заразили этот вид растений слизистой плесенью, у них стимулировалось производство фенольной кислоты, особенно вблизи места заражения:
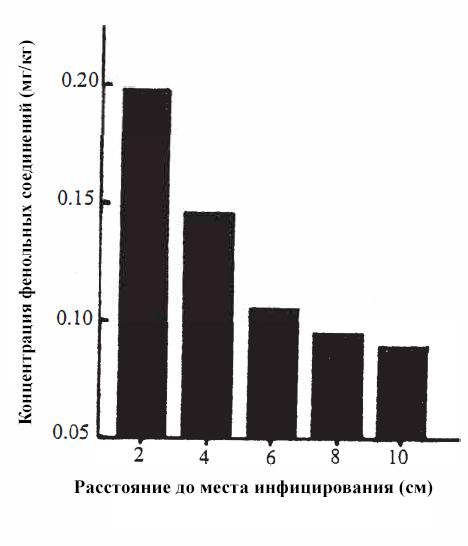
Концентрации фенольной кислоты вблизи места заражения листьев взморника морского (Zostera marina) из расчета сухого веса.
На расстоянии 2 см от очага поражения слизистой плесенью концентрация фенольной кислоты составляла около 0,2 мг/кг сухого веса, но на расстоянии 8 см от нее концентрация фенольной кислоты уменьшалась почти вдвое, примерно до 0,1 мг/кг. В частности, было показано, что концентрация кофеиновой кислоты в инфицированных листьях увеличивается примерно в 5 раз, тем самым достигая ингибирующих концентраций [29].

Взморник морской (Zostera marina). Z. marina, самая распространенная морская трава в Америке, растет большими подводными лугами. Когда Z. marina была намеренно заражена болезнетворной слизистой плесенью, растения синтезировали больше защитных фенольных кислот вокруг места заражения.
Растения, которым угрожают водоросли, могут увеличивать количество своей защитной фенольной кислоты. Большие паразитные водоросли (Caulerpa taxifolia), случайно завезенные в Средиземное море в 1984 году, вторглись на обширные луга из водорослей посидонии океанской (Posidonia oceanica) простирающиеся вдоль французского побережья. Водоросли прикрепляются к корневищу растения и впоследствии повреждают или убивают его. Исследователи [82] нашли, что площадь листа, занимаемая фенольными клетками у находящихся под угрозой исчезновения растений, составляла 43% , что почти вдвое больше, чем у растений из областей, которые не были заражены. Другое исследование выявило, что уровни фенольной кислоты (особенно феруловой кислоты) у находящихся под угрозой исчезновения растений были почти в два раза выше (641 мкг/г сухого веса), чем у растений из не зараженных областей (391 мкг/г сухого веса) [25].
8. Авто-ингибирование
Аллелопатическое автоингибирование, при котором растение подавляет рост своего собственного вида, было зарегистрировано у множества местных растений и сельскохозяйственных культур [1]. Например, в коре корней персикового дерева был обнаружен аллелохимический амигдалин (цианогенный гликозид). Почвенные бактерии расщепляют не обладающий подавляющим действием амигдалин на цианид, который в свою очередь сильно подавляет рост молодых персиковых деревьев [83]. Автоингибирование также было зарегистрировано у нескольких видов водорослей [84] и полупогружных водных растений [67]. Например, экстракты почвы, где растет тростник пестролистный (Phragmites karka) сильно тормозят прорастание семян этого вида.
Но почему растения выделяют соединения, которые подавляют их собственный вид? Один исследователь [73] объясняет автоингибирование тем, что оно, возможно, помогает растениям регулировать плотность собственной популяции. Зачастую автоингибирование подавляет семена и саженцы, но не взрослые растения. Ограничивая количество растений, особенно в стрессовых условиях, автоингибирование не уничтожает вид. Следовательно, автоингибирование может быть адаптивной стратегией, которая увеличивает выживаемость видов.
В.
У моих растений есть все: мощное освещение, грунт, представляющий собой идеальную смесь почвы, песка и вермикулита, подаю им C02, лью микроудобрения и вношу корневую подкормку. Растения прекрасно растут, а водоросли отсутствуют. Однако примерно через год растения начинают чахнуть и появляются водоросли. Это потому, что грунт становится все более анаэробным?
О.
Возможно, но анаэробный субстрат, вероятно, является следствием, а не причиной. Основная проблема заключается в том, что растения по какой-то причине перестали расти. Деградация грунта, которую вы видите, может быть связано с аллелопатией.
Многие виды растений выделяют аллелохимические вещества, и либо подавляют другие растения, либо сами себя («автоингибирование»). Аквариумный грунт на непроницаемом дне аквариума способствует постепенному накапливанию аллелохимических веществ. Более того, в вашем аквариуме с подачей СО2 и быстрым ростом растений, аллелохимические вещества могут накапливаться в грунте быстрее, чем разлагаться бактериями или связываться частицами почвы. Эти накопления автоингибирующей аллелохимии могут быть причиной, по которой ваш грунт "выдохся".
Аллелопатия у водорослей
Водоросли вырабатывют свои собственные аллелохимические вещества, некоторые из которых, вероятно, предназначены для конкуренции с другими водорослями, а некоторые - для отпугивания простейших и прочих травоядных, поедающих их. Предполагаемые целевые организмы часто трудно определить, из-за вторичных эффектов, изобилующих в водных экосистемах. Один исследователь (85) разочаровался в изучении аллелопатии водорослей в водной среде: "Аллелохимия настолько распространена в водных системах, что в нашей лаборатории, даже если мы специально пытаемся ее избежать, все равно находим ее везде, куда бы мы ни смотрели. Разобраться в этом - великая проблема."Другой исследователь [86] регулярно использовал нитчатые водоросли Pithophora, чтобы в его аквариумах не было других водорослей, особенно связанных с "цветением воды". В конце концов он создал 4 экспериментальных аквариума в которых содержались гуппи - одни с водорослями Pithophora, а другие без них. Несмотря на непрерывное освещение, аквариумы, содержащие Pithophora, оставались прозрачными в течение всех 4 недель, тогда как вода в аквариумах без Pithophora становилась зеленой через 7 дней. Рост зеленых водорослей, по-видимому, не имеет ничего общего с уровнями нитратов и фосфатов в воде:
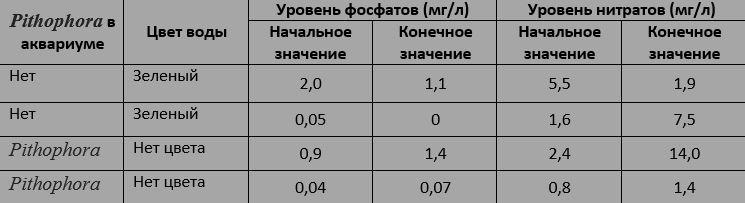
Влияние водорослей Pithophora на "цветение воды".
Водоросли как дырявые сосуды - выпускают из себя почти все вещества, которые вырабатывают, в том числе алеллохимические [87]. Один исследователь [85] обследовал более 200 различных "спариваний" водорослей из озера Коннектикут на предмет возможной аллелопатии. (Под "спариванием" здесь имеется ввиду воздействие термически обработанного фильтрата одного вида водорослей на другой вид водорослей.) Более чем две трети из этих 200 "спариваний" оказались аллелопатическими, так как фильтрат либо подавлял, либо стимулировал испытуемые виды. Более того, исследователь установил, что результаты лабораторных экспериментов соответствовали последовательности вспышек водорослей в самом озере. То есть водоросли, доминирующие в озере в течение одного сезона, выделяли

***
Комментарии: