
Ревизия эволюции, экологии и таксономии коридорасовых

Общее введение
На момент публикации книги "Идентификация панцирных сомов" (Фуллер и Эверс, 2005) ("Identifying Corydoradinae Catfishes", Fuller and Evers, 2005) было мало известно о взаимоотношениях между различными видами каллихтовых (Callichthyidae) и еще меньше о взаимоотношениях между коридорасовыми (Corydoradinae). Недавно ситуация изменилась благодаря публикации первой всеобъемлющей молекулярной филогении группы (Alexandrou et al., 2011). В этой статье мы обсуждаем достижения в филогенетике и экологии Corydoradinae и рассматриваем статус группы в целом с исторической точки зрения.Каллихтовые (Callichthyidae)
Семейство Callichthyidae (Bonaparte, 1838) - армированные сомы, широко распространенные в Южной Америке, определяемые наличием на теле двух продольных рядов боковых пластин, которые полностью покрывают бока, и двух или трех (основных) усиков на стыке губ по обе стороны рта (Gosline, 1940). Их название происходит от греческих слов kallis (красота) и ichthys (рыба). Семейство состоит из двух подсемейств: Callichthyinae (роды: Callichthys, Dianema, Hoplosternum, Lepthoplosternum и Megalechis) и Corydoradinae (Aspidoras, Scleromystax, Corydoras и Brochis). Corydoradinae составляют большинство (90% видов в пределах Callichthyidae) с более чем 170 описанными видами, плюс многие другие неописанные таксоны с номерами C или CW.
Прим. переводчика. Обратите внимание на разницу написания Callichthyidae и Callichthyinae. В биологической систематике иерархия обычно закреплена в окончаниях латинских названий. Так окончание -idae - означает семейство, а -inae - подсемейство.
Таксономическая история
Это, вероятно, очень старая группа. Найденные в формации Maiz Gordo Formation в Аргентине ископаемые коридорасы (Cockerell, 1925) датируются поздним палеоценом. Первый каллихтовый был описан Линнеем (Linnaeus, 1758) как Silurus callichthys (впоследствии переименованный в Callichthys callichthys). Впервые вид Corydoras был описан Lacepede в Histoire Naturelle des Poissons (Lacepede, 1803), где Corydoras geoffroyi был обозначен как типовой вид для нового рода. Впоследствии были установлены роды Brochis (Cope, 1872) и Aspidoras (Ihering, 1907). От Brochis (Cope, 1872) и Aspidoras (Ihering, 1907), Corydoras отличается сжатой с боков головой, короткими приротовыми усиками и единственной парой коротких подбородочных усиков на нижней губе. Род Brochis можно отличить по большому количеству мягких лучей спинного плавника (до 12 у Brochis против 6-8 у Corydoras и Aspidoras). Aspidoras отличается от Corydoras тем, что имеет короткий надзатылочный гребень, не разделяющий затылочные пластины (Nijssen, 1970). В 1952 году Hoedeman (Hoedeman, 1952) определил подсемейства Callichthyinae и Corydoradinae.
Таксономические связи Каллихтовых
До недавнего времени большинство таксономических и систематических исследований Callichthyidae были основаны на морфологических признаках и кладистическом анализе. Предпринималось много попыток классифицировать виды Corydoradinae по группам, которые отражают их эволюционную историю, используя различные методы, включая морфологию, цветовые схемы, число и характеристики хромосом, а в последнее время и генетические маркеры. Мы кратко рассмотрим работу, которая была проведена в каждой из этих областей.
Морфология
Первые комплексные исследования Corydoradinae были проведены Nijssen (1970) и Nijssen & Isbrucker (Nijssen, 1970; Nijssen and Isbrucker, 1980). В этих исследованиях использовались меристические подсчеты и морфологические соотношения, чтобы попытаться объединить виды в значимые группы. Однако, большинство видов, за некоторыми исключениями, пересекались по этим признакам, и их морфологический анализ при группировании таксонов был в основном неудачным (хотя и удачным с точки зрения характеристики видов). Из-за отсутствия различий Nijssen (1970), а затем Nijssen и Isbrucker (1980) предложили группировать таксоны по цветовым рисункам (обсуждается ниже). Следующая попытка навести порядок среди Corydoradinae была опубликована Штраусом (Strauss, 1985). Используя таблицы подсчетов и измерений Nijssen и Isbrucker (Nijssen and Isbrucker, 1980), Strauss выполнили многомерный статистический анализ, который показал, что изменения в морфологии являются результатом незначительных изменений в относительных скоростях роста различных структур тела (Strauss, 1985). Автор обнаружил, что четыре из пяти групп, предложенных Найссеном (1970), не могли быть должным образом дифференцированы из-за совпадения формы тела и меристики. Таким образом, результаты противоречили первоначально предложенным видовым группам Нейссена, которые в значительной степени опирались на соотношения и цветовые схемы.
Морфологический подход был использован в первом современном комплексном филогенетическом анализе, охватывающем все роды каллихтидов в рамках силуриформной структуры (Reis, 1998). Автор намеревался описать анатомию скелета Callichthyidae, чтобы изучить филогенетические взаимосвязи между видами и проверить монофилию семейства и его родов. Исследование подтвердило монофилию семейства Callichthyidae и разделение подсемейств Callichthyinae и Corydoradinae. В пределах Corydoradinae род Aspidoras был сестринской группой клада, образованной Corydoras и Brochis. Не было никаких признаков, которые подтверждали бы род Corydoras, тогда как монофилию рода Brochis подтверждали четыре наследуемых признака. Автор предоставляет ключ ко всем родам каллихтидов, основываясь на описанных морфологических признаках, и обсуждает группирование видов, как описано у Nijssen (1970) и Nijssen & Isbrucker (1980). Хотя анализ был надежным, но в нем отсутствовала таксономическая выборка в пределах Corydoradinae, особенно в родах Corydoras и Scleromystax.
В 2003 году Бритто опубликовал филогенетический анализ Corydoradinae, основанный на 83 морфологических признаках (Britto, 2003). Продвинувшись на шаг дальше, чем Рейс (1998), Бритто расширил число используемых морфологических признаков и общее таксономическое покрытие в пределах родов Corydoras, Scleromystax и Aspidoras. Одним из результатов этого исследования было предположение о немонофилетической природе Corydoras. Вместо того, чтобы Brochis и Corydoras сформировали клад с Aspidoras в качестве своей сестринской группы (Britto, 2003), Britto предложил клад, состоящий из Aspidoras и Scleromystax. Это привело к предложению двух племен в составе Corydoradinae: Aspidoradini и Corydoradini. Несмотря на тщательный анализ с использованием большого количества морфологических признаков, исследование не разрешило взаимоотношения внутри Corydoradinae и повысило вероятность того, что остеологические признаки могут не разрешать взаимоотношения в группе.
Цветовые схемы
С начала 19 века по настоящее время таксономические описания Corydoras были основаны главным образом на цветовых схемах. Они, как правило, изменчивы как внутри, так и между видами, что затрудняет их объективную количественную оценку (Nijssen, 1970). Это приводит как к смешению видов с одинаковым цветом, когда на самом деле они генетически, морфологически и географически различны, так и к разделению географических вариантов одного вида. В своей ревизии суринамских сомов рода Corydoras Nijssen (1970) предложил несколько групп видов, основанных на цветовой структуре, морфометрии и меристических признаках. Позднее они были уточнены Nijssen и Isbrucker (1980) до пяти групп. Однако эти группы не отражали филогенетическое родство между видами, что впоследствии было признано Isbrucker (Isbrucker, 2001). Таксономические проблемы также возникают при работе с миметическими видами, когда тип и материал паратипа относятся к разным линиям и они были объединены исключительно из-за их сходства в расцветке.
Хромосомы
Corydoradinae демонстрируют большие различия между видами в количестве хромосом, что делает их потенциально полезными признаками для видовой группировки. В первом исследовании сообщалось о кариотипическом и цитогенетическом разнообразии Corydoras (Scheel et al., 1972) и детальном подсчете хромосом у 14 видов Corydoras, число которых варьировалось от 44 у C. paleatus до 134 у C. aeneus. Авторы сравнили эти результаты с группами, предложенными Nijssen (1970), но не обнаружили очевидной связи между своими данными о хромосомах и группами Найссена. Дальнейшие исследования хромосом выявили 58 - 64 хромосомы в четырех популяциях C. aeneus (Oliveira et al., 1988) и 40, 42 и 44 хромосомы в трех популяциях C. nattereri (Oliveira et al., 1990). По сравнению с результатами, полученными Scheel et al. (Scheel et al., 1972), различия в данных C. aeneus указывали на существование возможной диплоидно-тетраплоидной системы в группе C. aeneus (58 - 64 против 134 хромосом). Дальнейшая работа охватывала виды Corydoras из более широкой географической области, выделяя характеристики хромосом и размеры генома для 11 видов из различных местностей всего южноамериканского континента (Oliveira et al., 1992). Это исследование увеличило общее количество видов Corydoras с известными кариотипами до 30.
Исследование заложило гипотетическую основу под эволюцию хромосом и размер генома в пределах группы. Впоследствии дополнительные виды каллихтидов были проанализированы Oliveira et al. с дополнительными гипотезами относительно эволюции генома внутри Callichthyidae в целом (Oliveira et al., 1993a; Oliveira et al., 1993b; Oliveira et al., 1993c). Эти исследования позволили группировать виды на основе количества хромосом, но взаимосвязи между группами и механизмы, приводящие к изменениям хромосом, были еще далеко не ясны.
Молекулярные филогенетические исследования
Первая молекулярная филогения семейства Callichthyidae была представлена Shimabukuro-Dias et al. (Shimabukuro-Dias et al., 2004). Авторы упорядочили митохондриальные гены 28 репрезентативных каллихтидов (роды: Corydoras (12), Aspidoras (3), Brochis (2), Dianema (2), Lepthoplosternum (2), Megalechis (2), Callichthys (2) и Hoplosternum (2). Также авторы пришли к выводу, что каллихтиды образуют монофилетическую совокупность, состоящую из двух природных групп: подсемейство Corydoradinae (роды: Aspidoras, Brochis и Corydoras) и подсемейство Callichthyinae (роды: Callichthys, Dianema, Hoplosternum, Lepthopslosternum и Megalechis). Эта молекулярная филогения противоречила ранее предложенным морфологическим отношениям. Добавление кариотипических данных привнесло уникальную филогенетическую перспективу, поддерживая базовые взаимоотношения диплоидных предков и монофилию групп полиплоидных видов в Corydoras.
В 2011 году Alexandrou et al. опубликовали первую всеобъемлющую молекулярную филогению Corydoradinae (Alexandrou et al., 2011). В этом исследовании для построения филогении группы использовались последовательности из множества генов с различными характеристиками. Они включают в себя митохондриальные и ядерные маркеры, которые широко используются в филогенетических исследованиях рыб и других организмов. Митохондриальные маркеры наследуются по материнской линии, так что все ее потомство (и самецы и самки) наследует исключительно ее митохондриальные гены. В филогенетической перспективе митохондриальные гены могут вводить в заблуждение в случаях гибридизации. Ядерные гены наследуются как от матери, так и от отца, и полезны для изучения гибридизации видов и групп. Используя комбинацию маркеров, можно придти к соглашению о наиболее вероятных отношениях между видами. Группа Alexandrou et al. (2011) в своем исследовании использовали последовательности из шести различных областей генома и все 435 отдельных таксона, включая несколько индивидуумов, когда это было возможно.
Филогения Alexandrou et al. (2011) определила девять основных линий Corydoradinae:
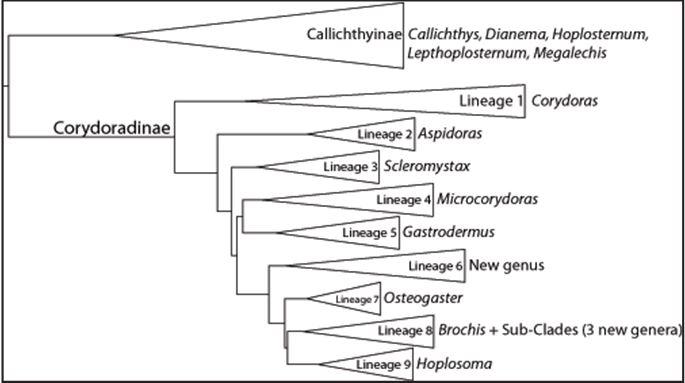
В порядке убывания времени, прошедшего с момента появления общего предка: Линия 1 (длинномордые виды, такие как C. fowleri), Линия 2 (Aspidoras), Линия 3 (Scleromystax), Линия 4 (карликовые + некоторые другие), Линия 5 (группа "elegans"), Линия 6 (C. paleatus и другие), Линия 7 (группа "aeneus"), Линия 8 (Brochis и такие виды, как C. haraldschultzi), Линия 9 (C. adolfoi и другие короткомордые виды). Эти результаты противоречат морфологической гипотезе, выдвинутой Britto(Britto, 2003), которая разделила племена Aspidoradini и Corydoradini на монофилетические сообщества. Поначалу точка зрения на племя Aspidoradini (Aspidoras и Scleromystax, Линии 2 и 3 соответственно) как родственной группы остальных Corydoradinae была отвергнута. Все молекулярные филогении, выявленные в этом исследовании, полностью поддерживают Линию 1 как родственную группу всех других Corydoradinae, а Aspidoradini (Линии 2 и 3) наследуют от Линии 1. В результате морфологической филогении (Britto, 2003) Brochis стал синонимом Corydoras, что полностью подтверждается исследованием Alexandrou et al. (2011) (однако в будущем должна быть ревизия, связанная с восстановлением рода Brochis). Существуют и другие различия с предполагаемой морфологической филогенией в структуре меньших кладов среди Corydoradinae, относительно размещения C. garbei, C. undulatus, C. gracilis, C. laveolus, C. ephippifer, C. hastatus, C. pygmaeus, C. maculifer, C. reticulatus, C. agassizi, C. ambiacus и C. ornatus. Базальное положение Линии 1, которое включает типовые виды Corydoras (C. geoffroy), означает, что род в настоящее время является парафилетическим по отношению к Aspidoradini.
Экология Corydoradinae
Мимикрия
Аквариумисты давно знают, что виды коридорасов, живущие вместе, имеют похожую расцветку. Что непонятно, так это то, как различные виды связаны генетически и как им удалось сосуществовать. Alexandrou et al. (2011) показали, что виды, живущие вместе и имеющие схожую окраску, почти всегда имеют отдаленные генетические связи и происходят из разных генетических линий. Среди некоторых известных исключений можно назвать S.barbatus, S.macropterus и C. ehrhardti, C. paleatus, которые являются членами одной и той же линии (Scleromystax и Линии 6 соответственно). Это позволило исключить ряд гипотез, которые могли вести к выводу, что симпатрические виды имеют схожий цветовой паттерн (экологическое видообразование, симпатрическое видообразование с помощью полиплоидии и возможность того, что симпатрические виды на самом деле были разными морфотипами одного и того же вида). Corydoradinae хорошо защищены броневыми пластинами, завершающимися острыми ядовитыми (токсичными) шипами, поэтому мимикрия Мюллера, по-видимому, является наиболее вероятным объяснением одинаковых цветовых узоров. Мимикрия Мюллера имеет место, когда два или более хорошо защищенных вида используют одну и ту же расцветку для взаимной выгоды. Эти взаимные отношения развиваются, потому что хищникам необходимо напасть на определенное количество особей, чтобы в итоге понять, что такой цветовой паттерн сигнализирует о том, что с его владельцем лучше не связываться. Разделяя окраску, виды распределяют цену обучения хищников. Это отличается от бейтсовской мимикрии, где незащищенные жертвы имитируют защищенные виды. Примерами бейтсовской мимикрии являются Corydoras diphyes и Otocinclus mimulus (Axenrot and Kullander, 2003) и Corydoras hastatus и различные Characiformes (например, Serrapinnus kriegi), где незащищенные Otocinclus и Characiformes имитируют защищенные виды Corydoras.
Исследование диеты с использованием стабильных изотопов
Непосредственное изучение рациона детритоядных рыб может быть не простой задачей из-за сложности выявления фрагментированных и частично переваренных продуктов, обнаруженных в желудках рыб. Alexandrou et al. (2011) в изучении различий в рационе диких пойманных Corydoradinae, использовали стабильные изотопы δ15N и δ13C, обнаруженные в мышечной ткани. Стабильные изотопы особенно полезны при исследовании диеты, так как они сохраняют сигнатуру в течение длительного периода времени, в отличие от содержимого желудка, которое показывает, только то, что выловленный экземпляр съел в день отлова.
Азот (δ15N) ступенчато увеличивается между трофическими уровнями; например, ткани плотоядных животных имеют более высокие значения δ15N, чем ткани травоядных, из-за их более высокого положения в пищевой сети (Peterson and Fry, 1987). Углерод (δ13C) может незначительно меняться в зависимости от трофического уровня, однако основным источником изменений являются источники первичной добычи, а значения δ13C, как правило, более полезны при определении мест питания(Rubenstein and Hobson, 2004; West et al., 2006). Результаты исследований Alexandrou at al. (2011) демонстрируют, что симпатрические виды из разных генетических линий питаются на разных трофических уровнях, и, поэтому виды с разной морфологией морды способны разделять ресурсы и не конкурировать за пищу. Эти различия определяют общественную структуру миметических групп (такое поведение можно наблюдать, когда длинномордые виды добывают корм, погружая головы глубоко в бентос, в то время как короткомордые виды добывают корм на поверхности субстрата):
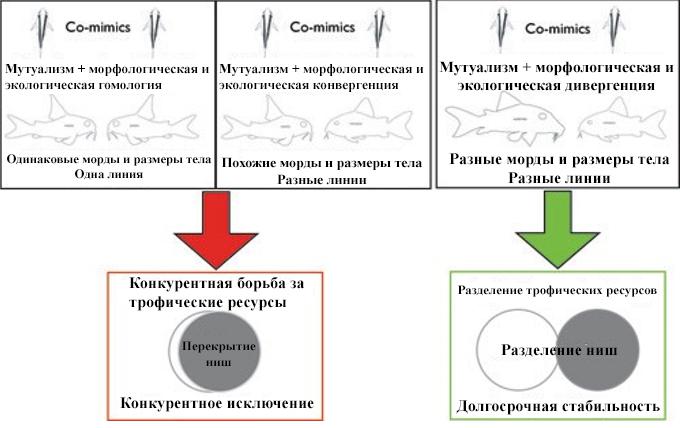
Некоторые представители Линии 4 (например, C. hastatus) не питаются бентосом, как их родственники, и занимают срединное положение в толще воды рек.
Morphology
Чтобы исследовать различия в морфологии морды и размерах тела у различных видов Corydoradinae, мы использовали геометрический морфометрический подход, основанный на ориентирах (Alexandrou et al., 2011). Вкратце, множественные ориентиры размещаются на идентифицируемых морфологических участках профиля рыбы.
Затем используется программное обеспечение для трансформации расстояний между ориентирами, что позволяет пользователю визуально исследовать и идентифицировать области, различающиеся у видов по форме. В исследовании Alexandrou et al. (2011) были сфотографированы и использованы для морфометрического анализа формы тела на основе цифровых ориентиров в общей сложности 200 имеющихся особей, представляющих более 120 различных видов (включая все миметические таксоны). Основные различия между видами, принадлежащими к различным линиям Corydoradinae, были выявлены в морфологии морды, положении глаза и высоте тела. Как упоминалось выше, различия в морфологии морды важны с точки зрения питания. Положение и размер глаза будут полезны для различения видов, принадлежащих к разным линиям, поскольку глаза видов в линиях 4 и 5 занимают сравнительно более низкую позицию по сравнению с другими линиями. Чувствительность геометрического морфометрического подхода достаточно велика, чтобы отнести любой вид к соотвествующей линейке с точностью до 99%. Это полезный подход, поскольку единственными данными, необходимыми для этих анализов, являются масштабные профильные изображения рыбы. Кроме того, морфологические различия внутри Corydoradinae сохраняются в пределах линии. Например, все виды Линии 9 - короткомордые, в то время как все виды Линии 1 - длинномордые. Даже в пределах линии 8, где присутствует некоторая морфологическая вариация (см. структуру подкласса ниже), виды могут быть идентифицированы по высоте их тела.
Систематические взаимосвязи и предложения по ревизии Corydoradinae (Hoedeman, 1952)
Из молекулярной филогении ясно, что Corydoradinae нуждаются в таксономической ревизии (Alexandrou et al., 2011). Необходимость такого пересмотра назрела уже давно (Isbri.cker, 2001), хотя только сейчас стали ясны генетические связи между видами, позволяющие группировать виды в монофилетические и филогенетически значимые группы. Здесь мы представляем предложения по ревизии Corydoradinae (как формально описанные, так и неописанные таксоны). Их следует рассматривать как общие предложения для будущего таксономического пересмотра и НЕ являющие собой официальную ревизию. Все виды должны продолжать называться Corydoras, Aspidoras или Scleromystax до тех пор, пока не будет опубликована тщательно рецензированная ревизия, включающая как морфологические, так и генетические данные.
Линия 1. Базальные "седлоносые" должны остаться под названием Corydoras, как их впервые описал Lacepede в 1803 году (Lacepede, 1803). C. geoffroy остается типичным видом для рода. Длиннорылые "седлоносые" типа Corydoras fowleri являются базовыми Corydoradinae. Виды этой линии включают в себя:
описанные виды: C. coriatae, C. fowleri, C. emiaquilus, C. treitlii, C. narcissus, C. serratus, C. simulatus, C. amapaensis, C. solox, C. cortesi, C. septentrionalis, C. stenocephalus, C. aurofrenatus, C. ellisae, C. blochi, C. pastazensis, C. acutus, C. areio, C. cervinus, C. geoffroy, C. heteromorphus, C. maculifer, C. negro, C. sarareensis, C. vittatus, C. ourastigma, C. oxyrhynchus, C. orcesi, C. saramaccensis
не описанные виды: c8, c16, c24, c28, c29, c38, c42, c47, c51, c53, c61, c63, c77, c78, c86, c92, c94, c95, c99, c109, c115, c116, c124, c127, c145, c146, c149, c153, cw5, cw11, cw12, cw17, cw53, cw55, cw59.
Линия 2. Эта группа останется Aspidoras (Ihering, 1907) с обозначенным типовым видом: A. rochai. Все известные Aspidoras принадлежат к линии 2 за исключением A. pauciradiatus. Кроме того, недавно описанные C. gladysae и C. petrarcini, по-видимому, тесно связаны с Aspidoras, однако у них отсутствуют определенные синапоморфии, которые определяют Aspidoras, поэтому потенциально требуется новое имя рода (Calvino and Alonso, 2009). Для того, чтобы лучше понять отношение этих видов к Aspidoras, необходимы генетические данные.
Описанные виды: A. albater, A. belenos, A. brunneus, A. carvalhoi, A. depinnai, A. eurycephalus, A. fuscoguttatus, A. lakoi, A. maculosus, A. menezesi, A. microgaleus, A. poecilius, A. psammatides, A. raimundi, A. rochai, A. spilotus, A. taurus, A. velites, A. virgulatus.
Не описанные виды: c35, c36, c37, c118, c119, c125, c158, cw52.
Линия 3. Все известные Scleromystax принадлежат к Линии 3, и, таким образом, эти группы должны оставаться Scleromystax (Gi.nther, 1864) с обозначенным типовым видом: S. barbatus.
Описанные виды: S. barbatus, S. macropterus, S. prionotus, S. kronei, S. salmacis, S. lacerdai.
Не описанные виды: c112, c113, cw38, cw42.
Линия 4. Эта линия включает в себя два вида карликовых, и поэтому мы предложили бы восстановление вышедшего из употребления Microcorydoras (Myers, 1953) с указанным типовым видом: C. hastatus. Не все виды в этой группе являются карликовыми как таковыми, но они тесно связаны и имеют схожую окраску на всем протяжении развития личинок.
Описанные виды: C. hastatus, C. pygmeaus, C. mamore, C. guapore, C. paucernA.
Линия 5. Линия 5 содержит виды, которые были известны как группа «elegans» Nijssen (Nijssen, 1970) с некоторыми дополнениями и исправлениями.
Ревизия может включать в себя восстановление рода Gastrodermus (Cope, 1878) с указанным типовым видом: C. elegans. C. gracilis является базальным видом в этой линии, и A. pauciradiatus также принадлежит к ней, а не к Aspidoras (Линия 2).
Описанные виды: C. gracilis, C. sp. A. paucirdiatus, C. nijsseni, C. bilineatus, C. elegans, C. nanus, C. napoensis, C. undulatus.
Не описанные виды: c41, c88, c89, c123, c126, c132, cw8, cw18, cw19, cw22, cw29, cw44, cw48, cw56.
Линия 6. Виды в этой группе всегда классифицировались под родом Corydoras, при этом в ней нет вышедших их употребления синонимичных общих названий. Таким образом, необходимо будет описать новый род с новым типовым видом.
Описанные виды: C. carlae, C. cochui, C. nattereri, C. potaroensis, C. diphyes, C. ehrhardti, C. micracanthus, C. paleatus, C. laveolus, C. reynoldsi, C. tukano, C. albolineatus, C. longipinnis, C. ortegai, C. steindachneri.
Не описанные виды: c7, c40, c73, c114, c144, cw3, cw24.
Линия 7. Эта линия включает в себя все виды из группы "Aeneus". Ревизия будет включать в себя восстановление рода Osteogaster (Cope, 1871) с указанием типового вида: C. eques. Наиболее базальными видами в этой группе являются C. melanotaenia и C. aeneus из Тринидада, встречающиеся в бассейне Ориноко. C. zygatus и C. rabauti состоят в этой группе и, по-видимому, более тесно связаны друг с другом, чем с другими видами в линии. Наиболее тесно связаны с этими видами C. aeneus spp. из бассейна Параны, которые первоначально были известны как C. macrosteus. Амазонские виды образуют группу внутри линии, причем C. aeneus из Суринама и Гайаны отделены от видов из Перу, где встречаются так называемые «лазерные» виды.
Описанные виды: C. rabauti, C. aeneus (spp.), C. eques, C. melanotaenia, C. zygatus, C. schultzei, C. venezuelanus.
Не описанные виды: cw7, cw9, cw10, cw14, cw16, cw23, cw26, cw41, cw43.
Линия 8. Эта линия включает в себя главным образом "промежуточных длинномордых" - длинномордых, но высокотелых, и также в нее входят Brochis, которые недавно были синонимом Corydoras (Britto, 2003). Ревизия должна восстановить название Brochis (Cope, 1871), с обозначенным типовым видом: B. splendens. Кроме того, еще три рода должны были бы быть названы для подкласса в пределах этой богатой разновидностями линии.
Описанные виды:
Подкласс 1: Brochis: B. britskii, B. multiradiatus, B. splendens.
Не описанные виды: cw34, cw35.
Подкласс 2: C. garbei, C. diluviatilis, C. filamentosus.
Не описанные виды: c57, c67.
Подкласс 3: C. latus, C. sodalis, C. reticulatus, C. geryi, C. pantanalensis.
Не описанные виды: c81, cw33.
Подкласс 4: C. crypticus, C. imitator, C. virginiae, C. amandajanea, C. condisciplus, C. ornatus, C. orphnopterus, C. pulcher, C. agassizii, C. ambiacus, C. crimmeni, C. delphax, C. ephippifer, C. incolicana, C. robustus, C. leopardus, C. gomezi, C. haraldschultzi, C. isbrueckeri, C. noelkempffi, C. pinheiroi, C. robinae, C. seussi, C. spectabilis, C. approuaguensis, C. filamentosus, C. sychri, C. melanistius, C. lamberti, C. spilurus, C. bifasciatus.
Не описанные виды: c9, c10, c13, c18, c34, c39, c49, c52, c66, c68, c71, c74, c75, c80, c87, c97, c98, c101, c102, c103, c110, c117, c122, c128, c130, c131, c135, c138, c140, c143, c152, c155, c156, c157, c159, cw2, cw6, cw13, cw20, cw25, cw40, cw57, cw58.
Линия 9. Виды Линии 9 - это классические виды с «короткой мордой», такие как C. adolfoi. Ревизия, вероятно, будет включать восстановление имени Hoplosoma (Agassiz, 1846), с указанием типового вида: C. punctatus. Это очень богатая видами линия, и многие виды выделились относительно недавно.
Описанные виды: C. boesemani, C. arcuatus, C. adolfoi, C. davidsandsi, C. duplicareus, C. melini, C. metae, C. panda, C. gossei, C. burgessi, C. griseus, C. oiapoquensis, C. baderi, C. concolor, C. axelrodi, C. armatus, C. atropersonatus, C. kanei, C. loretoensis, C. loxozonus, C. bicolor, C. brevirostris, C. evelynae, C. leucomelas, C. parallelus, C. schwartzi, C. habrosus, C. sterbai, C. trilineatus, C. araguaiaensis, C. bondi, C. breei, C. copei, C. coppenamensis, C. cruziensis, C. julii, C. multimaculatus, C. osteocarus, C. paragua, C. polystictus, C. punctatus, C. sipalwini, C. caudimaculatus, C. similis, C. weitzmani, C. urucu, C. xinguensis, C. sanchesi, C. surinamensis, C. boehlkei.
Не описанные виды: c3, C6, c14, c19, c21, c30, c33, c43, c44, c45, c48, c54, c62, c65, c76, c84, c85, c90, c91, c96, c100, c104, c120, c121, c129, c133, c134, c136, c137, c139, c141, c142, c147, c148, c150, c151, c154, cw1, cw4, cw15, cw21, cw27, cw28, cw30, cw31, cw32, cw36, cw37, cw39, cw45, cw46, cw47, cw49, cw50, cw51, cw54, cw60.
Благодарности: мы благодарим Juan Montoya-Burgos, Mark Sabaj Perez, Ian Fuller, Hans- Georg Evers, Mark Walters и Kim Mathiesen за предоставление образцов тканей и фотографии. Благодарим Claudio Oliveira, который помог провести полевые работы и внес вклад во все аспекты исследования. Мы также благодарим всех членов Группы по Изучению Сомов за их энтузиазм и поддержку.
Словарь:
Аллопатрический - встречается в отдельных непересекающихся географических зонах.
Бейтсовская мимикрия - мимикрия, в которой съедобное животное защищено своим сходством с ядовитым, которого избегают хищники.
Клад (клада) - группа организмов, которые, как полагают, эволюционировали от общего предка, согласно принципам кладистики.
Кладистика - это метод классификации животных и растений по пропорции измеримых характеристик, которые являются для них общими.
Конвергенция - тенденция к развитию внешне сходных характеристик в сходных условиях окружающей среды у неродственных животных и растений .
Цитогенетика - изучение наследования применительно к структуре и функциям хромосом.
Диплоид - содержащий два полных набора хромосом, по одному от каждого родителя.
Кариотип - количество и внешний вид хромосом в клеточных ядрах организма или вида.
Меристика - подсчет и измерение количественных характеристик рыб, таких как количество плавников или чешуек.
Монофилетический - происходящий от общего эволюционного предка или родовой группы.
Морфометрия - процесс измерения внешней формы и размеров рельефа, живых организмов или других объектов.
Мимикрия Мюллера - форма мимикрии, в которой два или более ядовитых животных проявляют схожий внешний вид в качестве общего защитного механизма. Теоретически, смысл заключается в том, что если хищник научится избегать одного из ядовитых видов, он также будет избегать весь мимический вид.
Разделение ниши - процесс, посредством которого естественный отбор приводит конкурирующие виды к различным моделям использования ресурсов или различным нишам.
Ниша - место, занимаемое видом в его экосистеме.
Парафилетический - происходящий от общего эволюционного предка или наследственной группы, но не включающий всех потомков.
Полифилетический - происходящий от нескольких общих эволюционных предков или наследственной группы и, следовательно, не подходит для размещения в одном и том же таксоне.
Полиплоид - организм или клетка, содержащая более двух гомологичных наборов хромосом.
Радиация - отклонение от центральной точки, в частности эволюция от предковой группы животных или растений в различные новые формы.
Стабильные изотопы - встречающиеся в природе стабильные формы элементов с разными ядерными массами, которые придают разнородные физические свойства, заставляющие такие изотопы вести себя по-разному в биогеохимических процессах.
Симпатический - встречающийся в одной и той же географической зоне; перекрытие в распределении.
Синапоморфия - обладание двумя организмами особенностью (не обязательно одинаковой у каждого), которая наследуется от характерной особенности организма, из которого они оба развились.
Трофический уровень - каждый из нескольких иерархических уровней экосистемы, включающий в себя организмы, которые выполняют одинаковую функцию в пищевой цепи и имеют одинаковые пищевые связи с основными источниками энергии (положение, которое организм занимает в пищевой цепи).
***
Комментарии (0):
Комментариев пока нет.